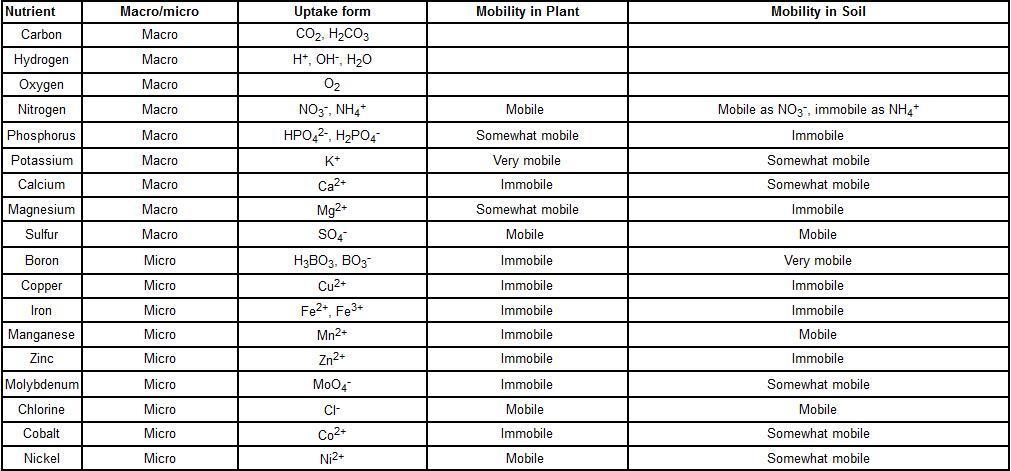
Ultraviolet_ "And God said, Let there be light, and there was light." Genesis 1:3
Inhale for 13s, Exhale for 13s,
Inhale for 8s, Exhale for 8s,
Inhale for 5s, Exhale for 5s,
Inhale for 3s, Exhale for 3s,
Inhale for 1s, Exhale for 1s,
Alignment. 1.618
More energy, more blueprint.
Electrical Conductivity refers to how easily a material or solution allows electrons or ions to flow continuously when an electrical potential is applied.
Electrochemical Reactance is the opposition to alternating current (AC) caused specifically by the temporary storage of energy in electric or magnetic fields, rather than energy being lost as heat. Reactance does not directly alter a material's intrinsic electrical conductivity. Instead, it dictates how the system stores and releases energy over time, which creates a temporary barrier to current flow in AC circuits. Together with resistance, reactance makes up total impedance. Electrical impedance (the combination of resistance and reactance) in the rhizosphere dictates how easily ions and water move into plant roots, directly impacting the Electron Transport Rate (ETR). High impedance restricts ion mobility, leading to nutrient deficiencies that decrease ETR and stunt overall plant growth.
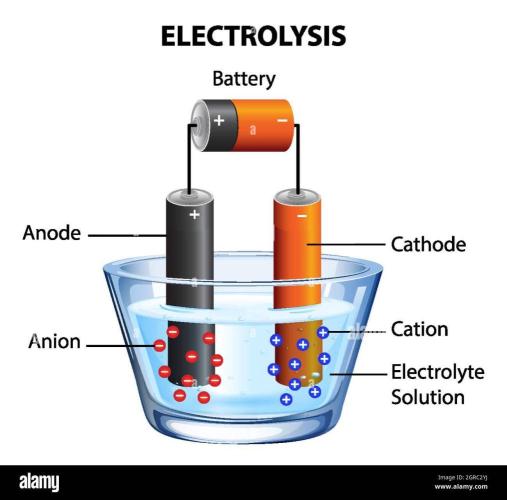
Using copper and zinc plates in the plant medium to form a natural earth battery to assist with Electrochemical Reactance. The moist soil acts as an electrolyte, while zinc serves as the anode and copper as the cathode. Zinc is a highly reactive metal and oxidizes, losing electrons into the soil. These electrons travel through an external wire to the copper plate. The moisture and dissolved salts in your plant medium allow charged ions to move freely between the plates, completing the circuit and generating a small direct current (usually between 0.8 and 1.1 volts). This micro-current subtly alters/the Electrical Conductivity (EC), allowing more effective breaking apart of chemical bonds in the soil (electrolysis), making soil nutrients like phosphorus, calcium, and potassium more accessible to roots. Buried the zinc plate on one side of the plant’s root zone and the copper plate on the opposite side. Ran an insulated copper wire above ground to connect the zinc plate to the copper plate, creating a closed loop. Make sure your soil remains moist and contains natural mineral salts; entirely distilled water or bone-dry medium will prevent ion exchange and block the current.
4 likes
comments
Share
Used method
Directly In Substrate
Germination Method
1
Week 1. Vegetation
2mo ago
1/18
20.32 cm
Height
18 hrs
Light Schedule
29 °C
Day Air Temp
6.5
pH
No Smell
Smell
640 PPM
TDS
50 %
Air Humidity
22 °C
Solution Temp
21 °C
Substrate Temp
24 °C
Night Air Temp
378.54 l
Pot Size
550 PPM
CO₂ Level
Ultraviolet_ "God is light, and in Him is no darkness at all." John 1:5.
Nirvana is interpreted as awakening from the illusion of separation and the limitations of time and space, effectively "seeing" the underlying code of existence. In Buddhism, Nirvana is described as the "unconditioned" (asankhata), a reality beyond dependent origination (cause and effect). This aligns with the concept of a fundamental, timeless "code" that exists before material manifestation. Some interpretations equate the stillness of Nirvana with the quantum vacuum, a state of "zero energy" which, however, holds unlimited potential for manifestation.
In quantum mechanics, a quantum system remains a "wave" of possibilities until observed. The "on-demand" aspect is often linked to the ability of a highly developed, enlightened consciousness (a "pure observer") to collapse this wave into a desired state, interacting directly with the underlying quantum reality. This view implies that by breaking free from the ego and mental conditioning, one gains mastery over the energetic, or vibrational, foundation of reality. The feeling of oneness in nirvana is explained through [quantum entanglement], suggesting all things are connected, and therefore, an enlightened mind can interact with any point in the universe. Traditionalists might argue that Nirvana is not about gaining control or having the power to "manifest on demand" (which can be a function of ego), but rather the total cessation of craving and the surrender of the ego, bringing an end to the desire to control reality.
The spiritual path is an evolution from being controlled by the "code" (samsara/suffering) to understanding the code and, eventually, becoming the author of one's own. With the awareness of how "code" functions, one takes full responsibility for their life, making conscious choices rather than reacting impulsively. Thoughts, words, and actions are purposefully aligned with higher values (truth, love, purpose), effectively rewriting one's destiny.
Humidity of the air dictates the moisture in the medium, which dictates the rate of pull from the terracotta stakes, 16 in total.
Soil-Plant-Atmosphere Continuum (SPAC): An ecological concept referring to the pathway of water moving from the soil, through the plant, and into the atmosphere.
Terracotta (unglazed, low-fired clay) is highly porous, acting as a breathable, permeable membrane that transports water based on the moisture gradient between the potting medium (soil) and the surrounding air. White terracotta and brown terracotta often have different pore sizes and overall porosity, primarily driven by differences in clay composition, impurities, and firing temperatures. While both are considered porous, brown terracotta often has higher iron content and impurities that behave as fluxes, affecting how the pores form during firing, while white terracotta is generally derived from more refined clays, with smaller pores making it more suited to pure water.
This mechanism is driven by capillary action and evaporation. When the surrounding soil is dry, the terracotta acts as a wick, pulling water out of the pot and into the soil. If the soil is very saturated, the terracotta absorbs water from the soil and allows it to evaporate from its outer surface, increasing the drying rate of the soil. Terracotta tends to keep the potting mix at an optimal saturation point, wicking up more water when the outside surface evaporates water into the air. The greater the difference in moisture between the soil and the outside air, the faster the water transfers through the ceramic. In low humidity and hot weather, the evaporation rate from the terracotta surface is high, creating a rapid drying effect. Newer terracotta pots often have a denser, lighter-toned structure that is less porous than traditional, red-orange terracotta, slowing down the moisture transfer rate. Over time, dissolved salts from fertilizer or tap water build up and block the pores, reducing the permeability of the clay. Water is all that is used or needed, thanks to the biochar and the massive storage bank of nutrients. Just waiting until something creeps up.
Clay pot irrigation (Ollas), an ancient farming practice, utilizes this property by burying unglazed jars in the soil, allowing water to slowly seep into the surrounding soil only when the soil dries out.
4 likes
comments
Share
2
Week 2. Vegetation
2mo ago
1/11
25.4 cm
Height
18 hrs
Light Schedule
29 °C
Day Air Temp
6.5
pH
No Smell
Smell
640 PPM
TDS
50 %
Air Humidity
22 °C
Solution Temp
21 °C
Substrate Temp
24 °C
Night Air Temp
378.54 l
Pot Size
700 PPM
CO₂ Level
Ultraviolet_ Not all minds think the same; it's a hard concept to grasp for one who has only ego. When someone operates solely from the ego, they tend to view their own perspective as the objective truth. To them, if someone else thinks differently, it is seen as a flaw, a lack of intelligence, or a personal affront, rather than simply a different, valid experience. The ego assumes that others are just extensions of itself. It cannot easily differentiate between "I think this" and "This is the universal truth". This is called the Mirror Fallacy. True understanding of another mind requires stepping out of oneself. The ego, by definition, is obsessed with self-preservation and being "right," which blocks empathy. Different perspectives are interpreted as attacks on the ego's stability. Instead of curiosity, it feels defensive. Acknowledging that others have entirely different internal worlds—with unique motivations, fears, and histories—requires dismantling the illusion that the world revolves around one’s own viewpoint. It’s a shift from "Why are they wrong?" to "How did they get to that perspective?"
Gospel of Thomas #17 "I shall give you what no eye has seen, what no ear has heard, and no hand has touched, and what has not come into the human heart." precedes saying #18, which questions the "beginning" and "end".
First, you need The Eyes to See, The Ears to Hear & The Mind of the Heart.
"Alpha to omega" refers to the first (Α/α) and last (Ω/ω) letters of the Greek alphabet, symbolizing "the beginning and the end". It represents totality, eternity, or the entirety of something.
Revelation 22:13, "I am the Alpha and the Omega, the First and the Last, the Beginning and the End" (NKJV, NIV).
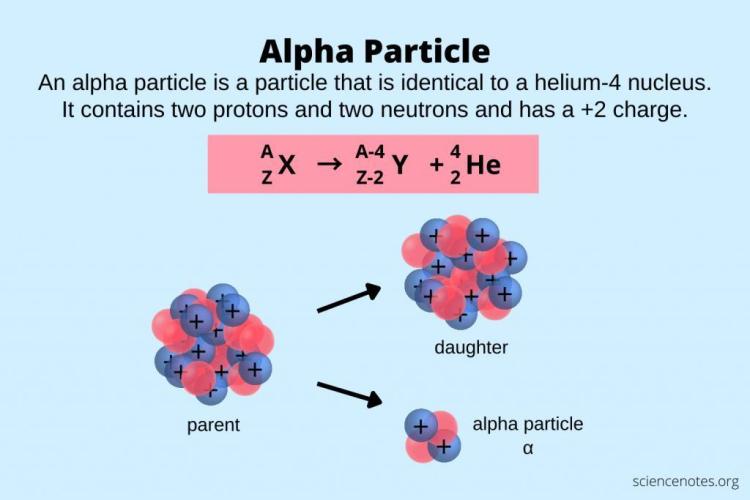
Both originate from Greek. Alpha (Α/α) is the first letter of the Greek alphabet (representing the beginning). Atom comes from atomos, meaning "indivisible," often seen as the foundational "first" particle of matter. In some esoteric and linguistic theories, the biblical name "Adam" (Hebrew for man/earth) is connected to "atom" because both represent the original, foundational unit (the first man vs. the first particle). Alpha Particles are particles emitted during radioactive decay, named "alpha" because they were the first type of nuclear radiation discovered. They are atomic nuclei consisting of two protons and two neutrons.
The study of the atom often deals with the "beginning" (alpha) of chemical reactions and fundamental building blocks. The terms are linked primarily as symbols for "the beginning" (Alpha), the "fundamental unit" (Atom), and the "original man" (Adam).
The uppercase symbol Omega (Ω/ω) is primarily used in electronics and physics to measure electrical resistance in ohms, representing how materials resist electron flow. It is also used in various fields to represent the density parameter in cosmology, solid angles in physics, and special constants or asymptotic growth rates in mathematics.
In quantum mechanics, "Omega" (Ω/ω) can refer to several concepts related to measuring or representing quantum coherence, generally categorized into dynamical, mathematical, or physical parameters:
Dynamical Coherence Measures
Omega Pattern (Superconducting Circuits)
Coupling Strength
Angular Frequency
Quantum Coherence Framework v24.0
Electron flow is not the same as electrical conductivity, but they are intimately related. Electron flow is the physical motion of electrons, while electrical conductivity is the material property that determines how easily that flow can happen.
Electron Flow (The "What"): The physical movement of negatively charged electrons through a conductor (usually from negative to positive). It is the act of carrying a charge.
Electrical Conductivity (The "How Well"): An intrinsic material property that measures how easily a material allows electrons (or other charge carriers) to move through it.
Electron Flow (The Action): The actual physical movement of electrons from one molecule (donor) to another (acceptor) in a pathway, such as the photosynthetic electron transport chain or cellular respiration. The current itself (dynamic electricity).
Electrical Conductivity (The Property): An intrinsic property of a material (like copper,glass,soil) that measures how easily it allows electric current (electron flow) to pass through it. A measure of ease. A high-conductivity material has low resistance and lets electrons move freely.
Electron transport rate (ETR), the rate at which electrons travel through the chain.
You can have Electron Flow occurring (electrons moving through the chain), but a very low Electron Transport Rate if the plant is in the shade. If light increases, the Electron Flow still follows the same path, but the Electron Transport Rate increases because more electrons are moving per second. In summary, ETR is a quantitative measure of electron flow. Think of "Electron Flow" as the movement itself, and "Electron Transport Rate" as the speed or quantity of that movement.
Photosystem II (PSII) is a critical multi-subunit protein complex in thylakoid membranes that initiates photosynthetic light reactions by splitting water and transferring electrons.
"IVM Matrix" or Isotropic Vector Matrix, a fundamental 3D grid of tetrahedra and octahedra described by R. Buckminster Fuller, representing an "isotropic" or uniform-distance, space-filling coordinate system. A structure where every vertex is equidistant from its neighbors, forming a network of triangles. Composed of 64 tetrahedra (often known as the 64-tetrahedron grid), 12 octahedra, and 6 octa-tetrahedra. It is a vector equilibrium, meaning it is the only geometric form where the distance from the center to its vertices is the same as the distance between its vertices. Considered a, if not the, fundamental geometry of space in synergetic geometry, often used to map quantum energy flows, such as in the 64-tetrahedron grid.
6 likes
comments
Share
3
Week 3. Vegetation
2mo ago
1/14
35.56 cm
Height
18 hrs
Light Schedule
29 °C
Day Air Temp
6.5
pH
No Smell
Smell
640 PPM
TDS
50 %
Air Humidity
22 °C
Solution Temp
21 °C
Substrate Temp
20 °C
Night Air Temp
378.54 l
Pot Size
800 PPM
CO₂ Level
Ultraviolet_ Topped once, turned off IR @ nights, slowed vertical growth back down, and took off both of the very lowest internodes on each plant.
Eisenia fetida
Stratiolaelaps scimitus
Armadillidium vulgare
Red wigglers (Eisenia fetida) are highly beneficial. They are considered an ideal choice for "no-till" or container-based organic growing because they live in the upper layers of soil, feeding on organic mulch rather than the plant's root system. Red wigglers accelerate the breakdown of organic amendments and produce high-quality, nutrient-dense worm castings directly in the root zone. Clover is another exceptional component of an organic rhizosphere, offering a sustainable, self-sustaining alternative to synthetic nitrogen fertilizers produced via the energy-intensive Haber-Bosch process. By forming a symbiotic relationship with Rhizobia bacteria, clover converts atmospheric nitrogen N2 into ammonium NH4, providing a steady, slow-release nutrient source that enhances soil health and reduces environmental impacts. Red clover offers superior nitrogen fixation and biomass production compared to white or yellow clover, making it the premier choice for maximum soil vitality, particularly for improving soil structure and providing a high-volume nitrogen credit for subsequent crops.
If it is fully functional and efficient soil, the rhizophagy cycle is superior long-term than any synthetic delivery when it comes to preventing deficiencies, not because it's "better," per se. The medium will require a very high CEC to make it to harvest without re-fertilization. The rhizosphere acts as a dynamic, interactive exchange where plants and soil microbes trade resources based on immediate needs. When a plant lacks a specific nutrient, it changes its physiology and releases specialized chemical cocktails—root exudates—into the surrounding soil. These exudates, which include sugars, amino acids, and organic acids, serve as a "shopping list" to attract specific microorganisms, which in turn return higher levels of desired nutrients. There is nothing in comparison when using synthetic delivery, which can cause plants to stop producing exudates, effectively "starving" the beneficial soil life, over time turning the soil barren and void of microbial life. Responsible use, applying the right amount at the right time, can minimize these negative effects. Relying solely on synthetic fertilizers without replenishing organic matter is what typically leads to exhausted soil. The use of synthetic fertilizers can utilize the Cation Exchange Capacity (CEC) of the soil, but without a robust rhizosphere and active microorganisms, the efficiency of this process is significantly reduced. This makes synthetic growing more difficult to prevent deficiencies overall compared to an efficient organic living soil with a robust rhizophagy cycle, as there is no "one size, fits all" when it comes to different nutrient profiles of strains/genetics, making it trickier to "guess" and prevent creeping deficiencies.
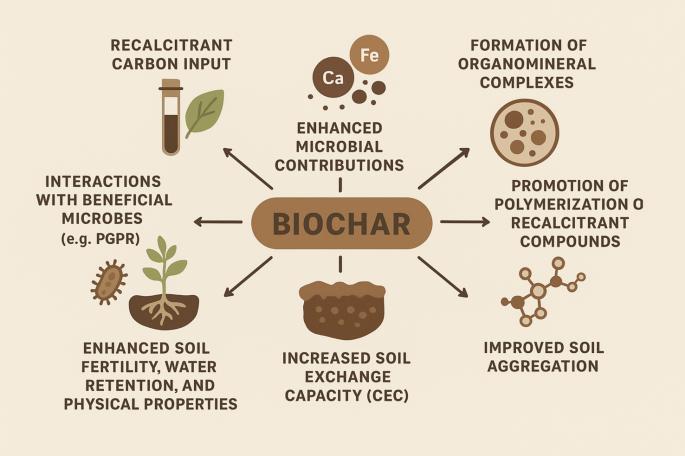
CEC does not contribute towards EC. Add more CEC using biochar, problem solved. If you keep pH between 6.3 and 6.7, hydrogen is exudated to cycle the medium's CEC for its needs. Keeping the pH between 6.3 and 6.7 creates an environment where plants release H+ to displace positively charged nutrients (like Ca2+, Mg2+, K+ held on soil particles or within artificial media this cycle through nutrients via the medium's Cation Exchange Capacity (CEC).
When biochar is first produced, it is highly porous and essentially empty. If you put this raw biochar directly into your soil, it will immediately act like a sponge, sucking up all the nutrients and water from your surrounding dirt. This can cause your plants to experience a temporary nutrient deficiency. Charging (or "inoculating") fixes this by pre-loading the biochar’s empty pores with nutrients and beneficial microbes. It fills the biochar's "storage" before it ever touches your garden. To charge your biochar, you soak it for a couple of weeks in nutrient-rich materials. Once charged, the biochar won't steal anything from your soil. Instead, it acts as a permanent, nutrient-rich reservoir that slowly releases fertilizer and water to your plants while providing a safe, protected home for soil microbes.
Microorganisms generate a stable potential of approximately 0.5 V EC. The rhizosphere creates its own food, similarly to chelation, using 1000's of varying combinations to create its own food.
Start to finish, just add water. Eventually, more materials will need to be added at the beginning of each new grow, but very attainable to go from seed to harvest without ever fertilizing.
ATP is important when it comes to biomass accumulation. Cellular root respiration and cellular respiration are essentially the same biological process, the breakdown of glucose to create usable energy (ATP) in the presence of oxygen, just taking place in different parts of the plant. Synthetic (salt-based) grows have significantly lower levels of total rhizosphere respiration, often referred to as root-zone activity, compared to organic living soil grows. While the plant roots themselves may respire in both systems, the surrounding soil ecosystem in a living soil setup is vastly more active, teeming with bacteria, fungi, and beneficial microorganisms.
2 pools of ATP, it won't double in growth buuuut, but improving root respiration by ensuring high oxygen in the soil is crucial. Good aeration ensures roots can fully utilize glucose to generate the ATP necessary for nutrient uptake, leading to healthier and more productive plants, even if growth isn't exactly doubled. The ATP created using root respiration is dedicated to rootzone growth; the ATP created using regular cellular respiration in a synthetic system would have to dedicate a lot of ATP to the roots when there is little or no root respiration. It's true that there is less of an initial ATP cost in breakdown when nutrients are already in their final form (synthetic), but you lose a solid chunk of ATP when the entire plant is reliant on cellular respiration alone; a large portion of ATP is dedicated to root zones for "forced" nutrient uptake rather than traded. Making it overall less efficient, even if the initial cost of breakdown is higher. Not sure if I butchered that, but one can hope it makes sense.
Oxygen is of critical importance when growing in living soil compared to synthetic soil because it supports the metabolic needs of the microbial, fungal, and insect ecosystem, rather than just the root respiration required by the plant itself. While synthetic grows can survive in lower-oxygen environments with precise mineral feeding, living soil systems rely on aerobic microbes to decompose organic matter (microbial mineralization) to create plant-available nutrients, which is an oxygen-intensive process. While a specific fair percentage is difficult to guess, my experience points to a massive, compound difference between the two methods and the amount of oxygen required. All the ATP spared is used on more biomass, not only that, but the extra root respiration can achieve a much higher CO2 compensation point naturally than you could with synthetic and atmospheric CO2 alone.
As a plant grows faster and increases in size, its demand for nutrients to support that growth increases, requiring a higher rate of nutrient uptake. As plants enter phases of rapid vegetative/floral growth, their metabolic demand for nutrients increases exponentially. Without a robust buffer zone—whether in the soil (cation exchange capacity) or in a hydroponic reservoir—deficiencies will occur rapidly because the instantaneous demand for specific nutrients can quickly exceed the rate of supply.
A growing body of evidence suggests that organic living soil provides superior long-term soil health and environmental benefits compared to synthetic fertilizers, which are often criticized for promoting a cycle of dependency and degradation. While synthetic fertilizers offer short-term convenience and high yields, they often come at the expense of long-term soil health, sustainability, and increased corporate control over growers/ farmers. Organic living soil, while slower and requiring more care to establish, creates a sustainable, resilient, and, ultimately, more fertile environment.
We don't really grow; we facilitate energy conversions, and energy is just numbers. Because the universe works the same way today as it did yesterday, there is a single, fundamental mathematical quantity that remains constant. We call this quantity energy. You cannot put "energy" under a microscope. You observe matter and forces (like heat, motion, or light), but energy is just a scalar number calculated to help predict how these things will change and interact. When an object falls, or when a battery powers your phone, matter shifts and changes form. Through it all, the universe ensures the "total score" of the numbers remains exactly the same.
95% to 97% of a plant’s dry matter consists of carbon, oxygen, and hydrogen. These three elements form the structural backbone of all plants. NPK & all the rest 3-5%.
Indigenous Amazonians created, or at least significantly enhanced, the fertile, dark soil known as Terra Preta de Índio (Portuguese for "Indian Black Earth") by incorporating biochar and other organic materials into the soil. This anthropogenic (human-made) soil technique, which dates back roughly 2,500 to 8,000 years, allowed ancient civilizations to flourish in regions with naturally poor, acidic, and nutrient-poor tropical soils.
7 likes
1 comment
Share
Used techniques
Topping
Technique
4
Week 4. Vegetation
1mo ago
1/10
48.26 cm
Height
18 hrs
Light Schedule
29 °C
Day Air Temp
6.5
pH
No Smell
Smell
60 %
Air Humidity
22 °C
Solution Temp
22 °C
Substrate Temp
20 °C
Night Air Temp
378.54 l
Pot Size
7.99 l
Watering Volume
800 PPM
CO₂ Level
Ultraviolet_ Remember that, however you are played, or by whom, your soul is in your keeping alone. Even though those who presume to play you be kings or men of power, when you stand before God, you cannot say, 'But I was told by others to do thus,' or that virtue was not convenient at the time. This will not suffice. Remember that.
Day:18
84°F and 65% RH (VPD) for the vegetative stage. Approximately 1.15kPa(assuming leaf temperature is about 2°F cooler than the air), which falls right into the ideal vegetative sweet spot (0.8kPa to 1.2kPa). At 1.15kPa, plants can draw water and nutrients efficiently without risking stress or wilting. It keeps the leaf pores (stomata) open, allowing for ideal carbon dioxide intake and maximizing vegetative growth. VPD is determined by the leaf's temperature, not just the ambient air. Because leaves usually run 1° to 3°F cooler than room air under bright grow lights, my actual VPD will be slightly lower, closer to the 1.0kPa mark. As she transitions from vegetative growth to flowering, one can gradually lower the humidity (to around 45–60%) and drop temperatures slightly to prevent disease from settling inside dense buds when they appear.
Night:6
At 70°F and 60% relative humidity, Vapor Pressure Deficit (VPD) is 0.86 kPa. This is right on the cusp of whats optimal for the vegetative stage. During the nighttime, plants generally close their stomata and undergo cellular respiration rather than photosynthesis. Transpiration slows to a near stop, making VPD less critical at night than during the day. However, maintaining a nighttime VPD between 0.8 and 1.0 kPa is highly beneficial in that it ensures the air is dry enough to prevent powdery mildew or bud rot, but moist enough to keep the plant from undergoing unnecessary stress. This range keeps the environment comfortable for cellular processes and prevents large atmospheric swings. Keeping it all flowing.
(Not pushing them yet, these are photoperiods)
The optimal soil (root zone) temperature for cellular root respiration and nutrient uptake in cannabis is between 68F & 72F This narrow range balances biological energy production (cellular respiration) with the dissolved oxygen levels in the soil, maximizing plant growth and health. Warmer soils hold significantly less dissolved oxygen. When soil temperature exceeds 74F oxygen depletion occurs, inhibiting cellular respiration almost entirely, At 68-72F root cells generate optimal adenosine triphosphate (ATP) via respiration to power root-tip elongation and the active transport of water and nutrients.
Too Hot (Above 78F) Root respiration increases, demanding more oxygen, while the water's oxygen-carrying capacity drops. This creates a prime environment for anaerobic pathogens and Pythium (root rot).
Too Cold (Below 60F) Root metabolism and cellular respiration slow to a crawl. This severely impairs nutrient and water absorption, leading to yellowing, wilting, and phosphorus deficiencies.
A lot depends on whether it's automatic or photoperiod; with photoperiod, there is not as much of a need to push "hard" as the real countdown only begins once the flower is initiated. Automatics, on the other hand, the chronological "clock" begins ticking the moment the seed germinates. It is of critical importance that the seedling growth gets off to the races, understanding that early growth is like compound interest, which will pay off come harvest. This reality is why getting autoflowers "off to the races" early on yields such exponential benefits. The "compound interest" is directly related to the surface area of the leaves. Larger, faster-growing seedlings process more light and build bigger root networks early on, which translates into an explosion of vertical and lateral growth during their short vegetative window.
The margins for error are so thin with autoflowers; this early-stage momentum depends on several critical practices. Seedlings exposed to increased atmospheric CO2 levels early in life will develop at an increased rate. To effectively "extend" or optimize the capacity of Photosystem II (PSII) for increased photosynthetic efficiency. In standard oxygenic photosynthesis, Photosystem II (PSII) is naturally limited to the red-light spectrum, peaking at 680nm. Extending its light-harvesting capacity past 700nm into the far-red region requires bypassing the natural limits of standard chlorophyll a. Adding 730 nm (far-red) LEDs alongside standard red/blue lights has been shown to increase canopy photosynthesis by 20–30% in several crops by acting synergistically with shorter wavelengths. However, the limitation is that excessive, pure IR/Far-red light (without accompanying red light) can trigger the "shade avoidance response," causing plants to grow tall, weak, and spindly rather than robust. Utilizing infrared light (specifically the 700-750 nm far-red range) is a viable method to boost photosynthetic efficiency. It acts as a bridge to allow PSII to utilize a broader spectrum of light, breaking the traditional 700 nm barrier. UVR8-mediated signaling (often in conjunction with CRY proteins) triggers protective mechanisms that maintain the stability of the photosynthetic apparatus (including LHCII and reaction center proteins), thus ensuring that the efficiency of Photosystem II remains higher in UV-B-exposed plants compared to plants lacking this receptor. ΦPSII indictates the rate of electron transfer from water to plastoquinone, which drives the production of ATP and NADPH. There is a close link between ΦPSII and the true rate of CO2 fixation (Φ*co2).
ETR stands for Electron Transport Rate. It measures the speed at which electrons are moved through the thylakoid membranes in a plant's chloroplasts during the light-dependent reactions of photosynthesis. Infrared light (particularly Near-Infrared or NIR) improves cellular energy by interacting directly with the electron transport chain (ETC) in mitochondria. This process boosts adenosine triphosphate production, which acts as a metabolic coefficient multiplier by accelerating enzyme activity dramatically.
Extend then multiply.
Far-Red photons interact with plant photoreceptors to accelerate the plant’s biological "clock" or trigger a shade-avoidance response. Autoflowers don't use the plant's biological clock, although the IR will initiate a shade avoidance and make them stretchy. You can just add equal measures of 660nm-680nm to negate the shade avoidance effect. Replacing nights' "darkness" with a combination of IR+ and 660nm. Because autoflowers don't require a dark period to flower, many growers just blast them with light. 18/6 24/0. However, this ignores the plant's metabolic rhythms, where daytime photosynthesis (light reactions) must be perfectly balanced with nighttime carbon fixation and assimilation (Calvin cycle) to avoid bottlenecking plant development. Cellular respiration is a 24/7 process, but it can only function while the plant has the free oxidative capacity to do so. A 100% photosynthetically active leaf cannot perform cellular respiration. The viral trend of defoliation of every leaf that isn't "getting enough light" is of great detriment overall, putting 100% of the cellular respiratory "workload" and responsibility on the 0/4/6 hours of darkness in sub-optimal conditions for enzymatic activity. Photosynthesis captures nearly 100% of the initial energy as carbon, while cellular respiration is the process that unlocks 90% of that captured energy into usable ATP so the plant can use it. Respiration is considered only roughly 30% to 40% efficient. It captures enough of the potential energy in glucose to synthesize around 30 to 38 ATP molecules per glucose molecule. The remaining 60% to 70% of the energy in the sugar is not captured in ATP; instead, it naturally escapes into the environment as heat, which helps regulate plant temperature. In plants, the primary enzymes of the Electron Transport Chain (ETC) and the ATP synthase complexes are typically adapted to function optimally in warmer temperatures (roughly 77°F to 95°F, depending on the specific plant strain). As temperatures rise within this physiological range, molecular collisions increase, speeding up respiration and ATP production. The cannabis plant has a branched respiratory pathway. During heat or cold stress, plants activate Alternative Oxidase (AOX). AOX burns sugars to dissipate energy as heat rather than coupling it to ATP production. This pathway actually functions optimally at elevated temperatures to help protect the cell from the damaging build-up of Reactive Oxygen Species (ROS) during heat stress. Enzyme activity generally scales with heat; there is a strict biological limit. If canopy temperatures in a grow room exceed 104°F, the enzymes and their supporting lipid membranes lose stability. Not saying you need to go crazy, just optimize nights the same as we optimize days.
Phosphorus is the driving force behind early seedling development. It acts as the "energy hub" of the plant, directly driving cell division, robust root growth, and the creation of DNA. Without an adequate, easily accessible supply early on, the plant's overall growth potential and final yield can suffer permanently.
6 likes
comments
Share
Used techniques
LST
Technique
5
Week 5. Vegetation
1mo ago
1/11
63.5 cm
Height
18 hrs
Light Schedule
28 °C
Day Air Temp
6.5
pH
No Smell
Smell
60 %
Air Humidity
22 °C
Solution Temp
22 °C
Substrate Temp
20 °C
Night Air Temp
378.54 l
Pot Size
11.36 l
Watering Volume
800 PPM
CO₂ Level
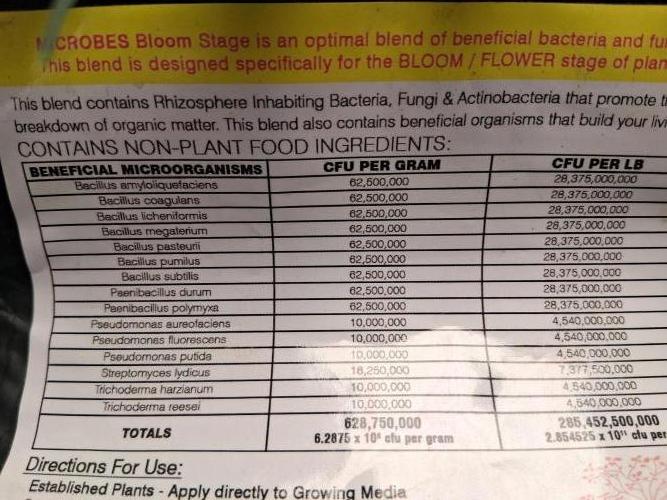
Nutrients 1
Microbes Bloom Stage
1.3 mll
Ultraviolet_ 18 hours in seconds, 60x60x18 = 64,800 seconds, now multiply by the 833μMol/s reading off the PAR metre. μMol/s (micromol) is the unit in which P.A.R. is expressed.
64,800x833=53,784,000μMol
53,784,000μMol = 54 Mol
54 DLI @ 800ppm, powerful combination. Pushing photosynthesis to peak capacity, accelerated growth and increased biomass. Applied net, stretching her around for a week. Maintaining a minimum air velocity of 0.3m/s within the inner canopy.
E=MC2 looks like a simple multiplication problem; it describes a fundamental physical truth: mass and energy are the same thing. The equation doesn't just calculate a value; it reveals that mass is effectively "congealed" energy. Energy is just numbers. Energy isn't a physical "substance" you can hold or touch. It is essentially an abstract, calculated number that we assign to a system to predict how it will change, interact, or move. A numerical label we attach to matter to track how it behaves. Because the universe runs on laws of symmetry (specifically, that the laws of physics don't change over time), a single global number must be conserved. We call that number "energy". The only difference is which side of the equation it's on.
How well a seedling grows is down to how much knowledge one can acquire to increase the level of conversion to occur. Applying knowledge effectively requires intuition, which comes from hands-on experience. A seasoned stoner learns to read subtle signs—like a slight change in leaf turgor (stiffness), subtle color shifts, or the specific texture of the soil—before a textbook diagnosis can be made. Ultimately, growing is the application of botanical science blended with active observation. Knowledge dictates your potential, but adaptability and attentiveness to the plant's immediate environment determine your results. The human eye is capable of distinguishing more shades of green than any other color in the visible spectrum.
1.618
Nature mathematically optimizes quantum energy transfer and light absorption efficiency within the photosynthetic machinery, as it naturally dictates energy scaling hierarchies and resonance dynamics. External vibration or electromagnetic wave that perfectly matches a plant's natural frequency directly influences plant growth. Low-frequency sound waves and targeted electromagnetic fields stimulate cellular processes and boost photosynthetic efficiency. An angular frequency aligned with the Golden Ratio is often used in wave mechanics and quantum physics to optimize energy transfer.
Alignment with what already exists.
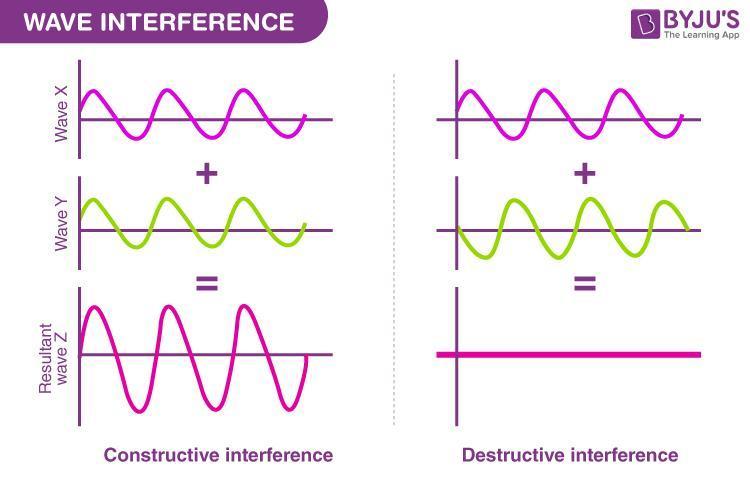
Law of nature, not my opinion: Constructive interference occurs when two or more overlapping waves meet in phase, meaning their crests and troughs perfectly align. Their amplitudes add together, creating a new, amplified wave with greater intensity than the individual components.
Constructive interference is the foundational mechanism that establishes a resonance hierarchy across physical, structural, and acoustic systems.
"Frequencies utilizing this ratio naturally minimize resistance (or impedance) and allow for highly efficient resonance, which can facilitate an increase in Electron Transfer Reactions (ETR) or energy pathways in molecular structures."
Quantum coherence in photosynthesis occurs when a photon of sunlight strikes a leaf; the energy it carries must travel to a reaction center to be converted into chemical energy. This process operates at nearly 100% efficiency. If the energy moved in a traditional "bunching" or random hopping manner, a large portion of it would be lost as heat. Instead, plants utilize quantum superposition. The energy particle (exciton) doesn't just take one path; it exists in a wave state and explores multiple pathways simultaneously. It essentially "chooses" the most efficient route to the reaction center simultaneously.
Research shows that molecular vibrations and the specific network arrangements of chlorophyll molecules (like the naturally evolved Chlorophyll A & B ratios) actively protect against energy overflow, optimizing light capture across different light intensities. Enzymes are the biological catalysts that speed up chemical reactions within a plant's cells, allowing them to grow, metabolize, and repair. Rather than relying solely on the classical kinetic energy of molecules colliding, plants use quantum tunneling. Subatomic particles like electrons and protons (hydrogen ions) can literally "teleport" through energy barriers that they normally wouldn’t have the energy to climb over. This makes vital metabolic reactions happen far faster than classical physics could ever explain!
Effective quantum efficiency of photosystem II, often denoted as ΦPSII, represents the proportion of light absorbed by Photosystem II (ΦPSII) that is actually used in photosynthetic electron transport. It is a key indicator of how efficiently a plant is using light for photosynthesis, as opposed to losing it as heat or fluorescence. ΦPSII (effective quantum yield of photosystem II) functions primarily as a "multiplier" (a coefficient of efficiency) rather than an additive factor when estimating the overall photosynthetic electron transport rate (ETR). Multipliers are considered far more beneficial than additions because they generate exponential growth, leverage existing resources to their full potential, and create sustainable, self-multiplying capacity, rather than just incremental, linear increases. This fascinating observation is rooted in the intersection of subatomic geometry, fractal scaling, and quantum dynamics. In specific molecular arrangements—such as in conjugated polymer networks or biomolecular architectures—the Golden Ratio (PHI) naturally dictates energy scaling hierarchies and resonance dynamics. Mathematically tied to the fine-structure constant, which defines the strength of the electromagnetic interaction. The Golden Ratio can be mapped geometrically as the Golden Angle (137.5 degrees) in atomic structures, linking the charge of the electron to fundamental quantum constants like Planck's constant.
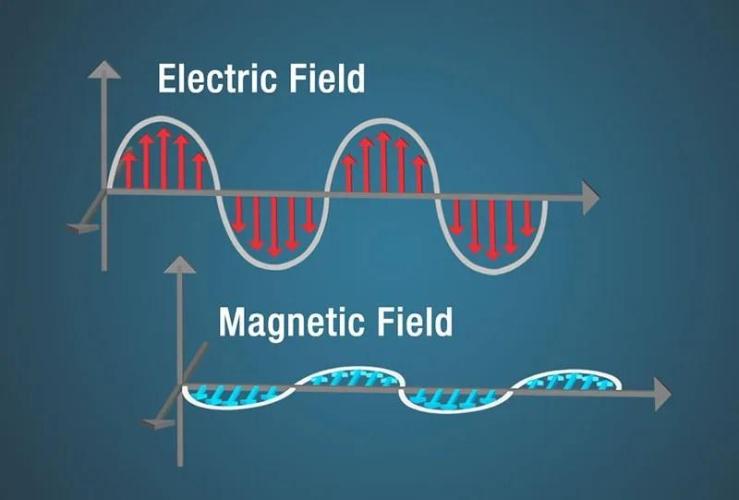
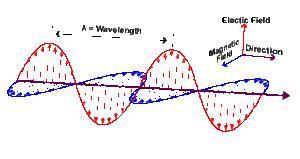
An electromagnetic wave is neither electric nor magnetic, but a combination of both. 137.5
The Golden Angle (137.5) This angle is derived from the Golden Ratio (1.618). It is the smaller of two angles created when a circle is divided such that the ratio of the arcs equals the Golden Ratio.
In quantum physics, Alpha (Α/α) is the approximate denominator of the fine-structure constant (often represented by the Greek letter Alpha (Α/α). It is a fundamental, dimensionless number that dictates how strongly charged particles (like electrons) interact with the electromagnetic field. Its value has puzzled physicists for over a century, as it bridges quantum mechanics, electromagnetism, and relativity.
In quantum mechanics, Omega (Ω/ω) usually represents angular frequency. When applied to "quantum coherence"—the ability of particles to exist in overlapping states without immediately degrading—is key to calculating how long these states last.
The Golden Angle, the value 137.5° is the geometric Golden Angle. It is derived from the Golden Ratio (1.618) and is famously responsible for the beautiful, mathematically optimized spiral patterns found in nature (such as sunflower seeds, pinecones, and leaf arrangements). Because of this, theoretical physicists and mathematicians have long speculated about why nature relies so heavily on the number 137 and the golden angle to govern everything from subatomic light interactions to the macroscopic geometry of the universe.
Stay with me, it's worth it.
7 likes
1 comment
Share
Used techniques
LST
Technique
Defoliation
Technique
6
Week 6. Vegetation
1mo ago
1/12
60.96 cm
Height
18 hrs
Light Schedule
31 °C
Day Air Temp
6.5
pH
No Smell
Smell
60 %
Air Humidity
22 °C
Solution Temp
22 °C
Substrate Temp
23 °C
Night Air Temp
378.54 l
Pot Size
7.57 l
Watering Volume
800 PPM
CO₂ Level
Nutrients 4
Microbes Bloom Stage
1.3 mll
RAW Yucca
0.7 mll
RAW Enzymes
0.7 mll
Ultraviolet_ The nighttime VPD does not need to mirror the daytime VPD. Daytime VPD dictates the pull of water and nutrients, while nighttime VPD acts mostly as a prevention tool. A high nighttime VPD increases the risk of the leaf temperature dropping below the dew point, which can trigger bud rot and powdery mildew.
Switched down to 12's hours of light mid-week, changed spectrum, increased light intensity from 840umol up to 1150umol at current height. Overnight from 800ppm to 1500ppm, morning compensation point (microorganisms) 46-47 days from germination, she fills the canopy herself, once the apical dominance is broken.
Measuring a plant's overnight CO2 emissions provides an accurate estimate of its dark respiration rate. Because photosynthesis stops in the dark, you are isolating the respiration process, which allows you to gauge how much stored energy (glucose) the plant has consumed and estimate the scale of oxidative phosphorylation. Oxidative phosphorylation is the final stage of respiration that generates the bulk of the plant's ATP (90%) and relies directly on the oxidation of these respiratory substrates NADH and FADH2 along with the consumption of oxygen.
From a thermodynamic standpoint. Growth is an energy-capturing process, and the rate of that growth is bound by the available free energy (Gibbs free energy) and the First Law of Thermodynamics. While the ceiling or upper limit is dictated by free energy (such as photosynthetically active radiation), the actual amount of growth relies on how the plant balances that energy with other limiting factors. These are often described as the nine cardinal parameters of plant growth. 4 Above, 5 Below. If any one of the 9 becomes bottlenecked, the entire plant's cycle is restricted.
Operating an 80F+ environment at night to force rapid carbon conversion comes with major drawbacks, as the biochemical processes work differently than the deductive logic suggests. While raising nighttime temperatures to 80F indeed accelerates respiration and speeds up the conversion of captured sugars (sink activity), doing so also radically increases the plant's overall metabolic baseline. If the plant's metabolic rate is artificially forced too high via heat, it can actually "burn" through more energy than it managed to assimilate during the day. This leads to carbohydrate starvation, stretching, and a net loss in final biomass yield.
400 ppm is near the standard ambient level; the plant's stomatal intake is the primary limiting factor, not the dark-reaction enzymes. To push 45 DLI without burning out the plant. Trying to force the conversion of a massive daylight DLI in a compressed time frame (12 hours) becomes highly inefficient because the Rubisco enzyme simply hits a saturation limit. To successfully convert a 45 DLI into dense, productive mass, the ambient CO2 generally needs to be elevated to the 1000 to 1200ppm range. This creates a steeper concentration gradient, driving the stomata to inhale CO2 fast enough to match the high photon energy.
It's not all about the amount of light, but the ratio too, as this will dictate growth through the ratio of phytohormones. In order for correct bud development, there needs to be a correct ratio of RGB. Different wavelengths have different penetration depths. When one grows using top-down lighting, only the entire canopy is limited to 2-3 layers of leaf, meaning there will only be correct bud development in those layers, regardless of getting 45DLI.
The biomass potential of a plant is linked to root mass. Generally, when a plant reaches its maximum biomass, you can help to chop off parts of the plant that are in less than efficient areas of the plant (low light) so that it can create new biomass growing towards the light.
Strength is the maximum potential, and power is the rate of conversion. You can have the biggest veg period of 18 weeks, and it means nothing, as soon as you start flowering, the chronological clock starts ticking, the only metric that matters to bud size is how much energy you convert each cycle, not by how long it took you to build the framework, it helps a lot nonetheless.
Not saying anyone should not defoliate for a reason, only that you should have one, and at the right time. Don't defoliate 30+% on autoflowers or 4 weeks into the flower period and expect an increase in yields; it doesn't work like that. There is room for dictating growth patterns and clearing out overcrowded nodes, but it needs to be done in veg because once that timer starts and buds start growing, it's all just energy conversion. One barely needs to defoliate at all in a 4x4 because with side lighting, turning a 2d canopy penetration into a 3d, even lower buds are 90% the quality and density of top ones. The rate of photosynthesis and the ultimate density of lower buds aren't just about the sheer number of photons PPFD. The specific ratio of R:G:B dictates canopy penetration and drives different photochemical reactions. The Electron Transport Rate (ETR) measures the speed at which electrons are driven through Photosystem II (PSII) during photosynthesis. The ratio of Red, Green, and Blue (RGB) light heavily dictates this rate.
Plant leaves continuously perform cellular respiration regardless of the time of day, using energy and oxygen to fuel essential metabolic maintenance. If you over-defoliate, the remaining canopy may be unable to produce enough net sugars during the day to offset the constant respiratory demands of the plant. Must balance fixation with assimilation; there's no point in capturing 45 DLI if you only convert 20% every cycle due to an extreme lack of respiratory capacity to perform cellular oxidative phosphorylation.
You can have a 4x4 canopy or a 4x4x4 canopy, yes, we know that side lights are not as effective at absorption from the sides or underneath, but it's not about DLI, it's never been just about efficiency, it's about the penetration ratios of RGB that drive ETR of/photosynthesis and trigger correct bud development. The size of each bud is its own ability to perform the ETR required for its own personal growth, and bud development is dictated by the ratio of RGB. It drives localised growth and acts as a regulatory switch for that development. Turgor pressure is another very important factor in understanding if you want big buds, for it is the "steam engine" that dictates the rate of bud expansion. Simply, a lot harder to achieve metabolically at ambient 75F than at say 86F
Because buds have less chlorophyll, they do not suffer from the same photosynthetic shutdown that over-exposed, light-stressed leaves do. They can soak up direct light energy to swell in density and size. Their tolerance to intense light is heavily limited by the temperature and humidity, but if you can control those temps and keep the rot away, buds have a much, much higher tolerance to high light than leaves. Beneficial to hammer with high light before trichomes appear. Balancing this with trichome maturity is key for rich terpene and flavonoid profiles, want it just right, somewhere in the middle, not too much, not too little. Find cannabis plants can defoliate themselves come harvest, given the right signals. Every last ounce of potential is recycled into buds by the plant itself (senseceance), given you can keep the level of conversion high enough to prompt a need to do so.
Get the canopy @ optimal PPFD range, 45-55DLI, then let the plant "stretch" the stems into a "PPFD range much higher, one that leaves don't like to grow in, but buds thrive in. What is optimal for a bud is different from what is optimal for a leaf photosynthetically. Genes provide the blueprint, but the environment dictates how, when, and if those genes are expressed. Must first signal the condition to increase the expression you want to exist through stress and response, cause and effect. A well-buffered CEC medium prevents extreme nutrient swings, allowing plants to maximise their dedicated genetic expression.
A plant is either genetically expressing "growing" or "recycling" genes based on its nutrient starvation level in the medium. Constantly toggling between "growing" and "recycling" hormonal states creates a futile cycle that wastes valuable metabolic energy. Plants rely on sophisticated biochemical switches to manage this trade-off and prevent rapid fluctuations that disrupt that balance.
This energy inefficiency is a recognised biological challenge. Plants avoid this costly "flip-flopping" by using hierarchical master regulators (like the TOR and SnRK1 protein kinases) that act as strict molecular switches. These networks enforce cellular commitment to either growth or survival, preventing mixed signals.
This is something that was missing from previous grows.
Under nutrient-rich conditions, TOR promotes protein synthesis, cell division, and structural expansion.
Under starvation, TOR is inhibited, and SnRK1 is activated. This triggers autophagy—where the plant breaks down old macromolecules and organelles to scavenge and reallocate essential nutrients to critical sinks.
"What's the point in flushing?"
The core idea behind a PK booster is to deliver a massive, concentrated surge of P&K exactly when buds are swelling in conjunction with a N starvation. Because these are short, targeted windows, the nutrients must be highly bioavailable so the plant can process them immediately. As soon as you go "organic," that's out the window. Much slower release, uncontrolled, very difficult to "spike". to cause the ratio that will initiate a response.
High-volume PK spikes rely strictly on the immediate uptake capabilities of mineral fertilisers. Making it far less efficient in organic/living soil setups.
When you use organic nutrients, it changes the dynamic with which the plant delivers and trades its nutrients; organic is always releasing new nutrients into the immediate EC. This prevents a lot of autophagic responses from occurring due to a constant stream of new nutrients into the immediate medium's EC. This can prevent nutrient starvation from being signalled.
PK boost is essentially just N starvation, triggering an autophagic response. Concentrated ratio of P&K while tapering off the Nitrogen base. To the plant, the sudden drop in Nitrogen registers as a severe environmental stressor—essentially, the beginning of starvation protocols. She aggressively strips nutrients and proteins from older leaves and vegetative structures and shuttles them directly to the developing flowers and fruit. Ta daaa. Call it a PK booster and sell it. Nothing to do with the P and K itself, it's the ratio immediately available in the medium triggering a nutrient recycling mechanism within the plant itself; all the "booster" sells is the trigger to the signal.
PK BOOST with 50% ammoniacal N signals floral maturation.
PK BOOST with N starvation signals nutrient recycling/sinking.
Very difficult to initiate a response when organic nutes are doing their thing. It takes 4x5x more water significantly to leach or wash ammonia out than it does nitrates. This can prevent triggering N starvation from having its normal impact.
Manipulating the C:N ratio in the medium. One autophagic response has multiple potential signal triggers. Nutrient starvation is not an option.
4 likes
comments
Share
Used techniques
LST
Technique
Defoliation
Technique
7
Week 7. Vegetation
20d ago
1/17
88.9 cm
Height
12 hrs
Light Schedule
31 °C
Day Air Temp
6.4
pH
No Smell
Smell
55 %
Air Humidity
22 °C
Solution Temp
22 °C
Substrate Temp
23 °C
Night Air Temp
378.54 l
Pot Size
7.57 l
Watering Volume
1400 PPM
CO₂ Level
Ultraviolet_ Wakíŋyaŋ, I am who I am, the salt of the earth.
Thunderbird is an allegory; his conflicts with other forces in nature are then an attempt to allegorize relationships observed in the natural order, such as the changing of the weather. He is essentially an attempt to represent the patterns of activity of a powerful, mysterious force in a way that can be understood simply and easily – sort of the way in which a weather map functions today.
Moving from18x60x60 = 64,800 seconds in 18 hours.
64800x860(ppfd) = 55,728,000 umol per daylight.
Into Flower
12x60x60 = 43,200 seconds in 12 hours.
43200x1145(ppfd) = 49,464,000 umol per daylight.
It's asking a lot of Rubisco regeneration to maintain 50 DLI in the 12 instead of 18. Raised the ambient CO2 to 1200 to 1500 ppm to achieve efficient gas exchange.
Not particularly recommended, but adding sugar to an indoor growing medium is a highly effective way to stimulate microbial activity, which rapidly breaks down the sugars and releases CO2 through cellular respiration. You can safely capture this CO2 to fertilize indoor crops and boost photosynthesis. While this process works, the setup requires precise understanding and management to avoid common indoor growing hazards.
The plant Carbon to Nitrogen C:N ratio defines the balance between structural carbon (sugars/cellulose) and nitrogen (proteins/enzymes). It acts as a master regulator of plant health, growth, and metabolism.
Rubisco (Ribulose-1,5-bisphosphate carboxylase/oxygenase) is the engine of photosynthesis responsible for fixing atmospheric CO2 into sugars. It is intimately tied to the C:N ratio for three primary reasons. It is the Plant’s Biggest Nitrogen Sink, Drives the Carbon Side, and it is the Nitrogen Control Knob. Understanding this relationship allows you to predict how plants respond to environmental stress or fertilizer.
Rubisco acts as the primary storage sink for leaf nitrogen, accounting for up to 30% to 50% of a C3 plant's soluble protein. Deep Green Leaves signal a rich abundance of both chlorophyll and Rubisco proteins. The plant possesses the heavy enzymatic machinery required to handle 1145 PPFD. Pale or yellowing leaves indicate a nitrogen deficiency. The plant is actively breaking down its own Rubisco to salvage nitrogen for newer growth, drastically reducing its light-tolerance threshold. Subtle difference, but understanding is important in order to be able to judge when to dial light intensity up and light intensity down, when to push, and when to back off. An extra dose of magnesium is vital if a plant is going to push through the growing pains of high-intensity lighting.
Foliar application of magnesium is an excellent and rapid way to assist with Rubisco regeneration within a plant, so long as it is applied correctly.
Spray strictly in the early morning or late evening, mixing your magnesium with a little fulvic acid or chelator, but only when she gets a little limey on top.
This, for me, is the experience of growing, akin to "riding the surf" maintaining efficient Rubisco regeneration through visual identification of the shade of green. Surf a razor-thin wave when balancing light intensity, nutrient availability, and transpiration to maximize Rubisco enzyme efficiency. Keeping the Calvin cycle fully charged without tipping into nutrient toxicity, light stress, or the dreaded chlorosis requires paying close attention to the visual cues the plant provides. By monitoring these subtle shifts in color, turgor pressure, and leaf posture, you adjust your environmental controls and surf that exact razor-thin wave.
Nute recycling acts as the vital execution mechanism for autophagy, which defines senescence. Natural senescence is a genetically programmed developmental stage aimed at nutrient recycling, whereas triggered autophagy is a rapid survival response activated by environmental stress. While both processes utilize the vacuole to break down cellular material, their triggers, selectivity, and overall goals are entirely different. Cannabis plant senescence is not separate from nutrient recycling protocols; rather, nutrient recycling is the primary physiological purpose of senescence, and autophagy serves as the core switch mechanism executing both processes.
Takes about 24 to 48 hours to notice visible changes once the signals have initiated the autophagic response. Not too late at all. A little bit of fade from senescence 2 weeks from harvest is normal and genetically expected. Send the C:N 32:1 signal 1 week from harvest for the best effect in your organic grow.
Understanding what makes leaves fade is not always senescence, but also strongly linked to Rubisco regeneration. That's a whole other subject. Vital to understand the differences if you want a correct diagnosis and to transition from hobby grower to master stoner, differentiating between a true genetic fade and a decline in photosynthetic proteins.
Nitrate is nitrate, whether it oxidizes or not is not up for debate. If it's not sunk by the plant you are smoking some if not all of, it's regardless of what your feelings are on the matter. Senescence is highly critical. It is the natural end-of-life stage where the plant redirects energy to ripen flowers. Properly managed, it breaks down harsh chlorophyll, allowing the terpenes (which provide taste and aroma) to peak. Harvesting outside this window leads to an "unripe" or degraded flavor comparable to going without.
To initiate the response you seek, you can trigger it multiple ways, when growing synthetically its triggered by nutrient starvation, generally when the entire medium is flushed. This is more to do with N starvation than being entirely empty. Nonetheless. PK boosters are N starvation through maximizing P and K. (Generally only works for synthetic grows)
Normally, a medium only holds 10-30% of its nitrogen as ammoniacal ta part boosts this to 50% as it triggers the "ripen" signal, but you don't want to keep ammoniacal above 30% for more than 7-10 days if you can help it. Its a trigger mechanism no more.
PK BOOST with 50% ammoniacal N signals floral maturation.
PK BOOST with N starvation signals nutrient recycling/sinking.
Because you are using organic nutes and you want to maintain the rhizosphere, what you want to do is add carbon in the form of sugars (powdered molasses). It's almost impossible to empty a medium enough when microorganisms are constantly releasing nutrients into the direct EC. Very difficult to initiate starvation responses with ammoniacal nitrogen. Manipulating the C:N ratio is the key to triggering an autophagic response and resulting nutrient recycling in the last days using organic nutes and without having to flush.
Generally not recommended for new growers. So do what you want. But if you don't trigger the plant to dump its nitrates into root zones, you will smoke nitrates as NO3- does not oxidize during the dry and cure no matter what you do or how long you dry or cure. Doesn't matter what anyone "feels" about it, how many grows they had with no fade.
"Clover steals valuable nutrients."
Crop and drop the clover come flipping to flower, its benefit comes from creating an airy and porous rootzone. I don't need to crop and drop once the plant fills the canopy, she blotches out the light, and the clovers die. This is the nitrogen the microorganisms use to convert carbon for respiration throughout the flowering stage.
5 likes
comments
Share
8
Week 8. Flowering
16d ago
1/21
114.3 cm
Height
12 hrs
Light Schedule
29 °C
Day Air Temp
6.4
pH
No Smell
Smell
60 %
Air Humidity
22 °C
Solution Temp
22 °C
Substrate Temp
23 °C
Night Air Temp
378.54 l
Pot Size
7.57 l
Watering Volume
1400 PPM
CO₂ Level
Ultraviolet_ 1.1 kPa VPD Days, 85F 60RH%
0.97 kPa VPD Nights 75F 55RH%
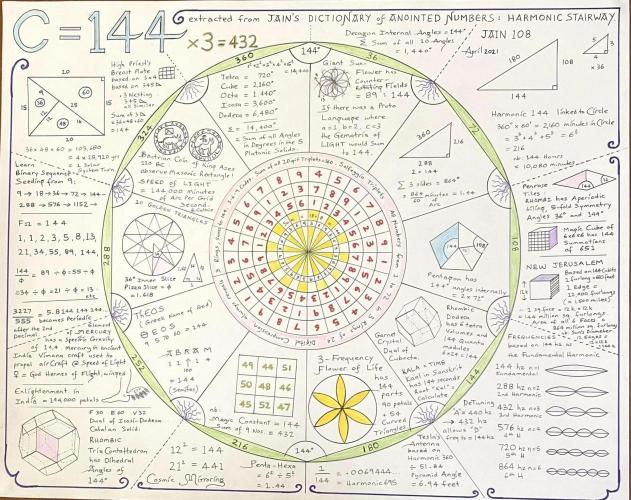
In 1200 AD an Italian noticed that plants grow from the ground in a certain way. The man was called Leonardo Pisano and he lived in Pisa. Today though, he is remembered at Leonardo Fibonacci from filius Bonacci, which means “the son of Bonaccio”. Simply by observing nature, he rediscovered the long-lost creation sequence:
1,1,2,3,5,8,13,21,34,55,89,144…
Pythagoras called this God BREATHING ON THE NUMBERS, but today it is simply referred to as the Fibonacci sequence. This series is neither arithmetic (based on the addition of numbers by a constant: 1, 2, 3, 4…) nor geometric (based on the multiplication of numbers by a constant: 2, 4, 6, 8…) but instead recursive. This sequence is based on the addition of two adjacent numbers to produce a third: 1, 1, 2, 3, 5, 8, 13…If we wished to express this sequence as an equation it would look like:
x + 1 = x²
Solving for x using the quadratic equation we get x = 1.618. When using this number we denote it as Φ and refer to it as Phi in commemoration of the Greek sculptor Phidias. We call this relationship the GOLDEN RATIO or 1:1.618.
But Leonardo Pisano did much more than rediscover the Fibonacci sequence—he brought “Indian numbers” to Europe. The familiar digits of 9, 8, 7, 6, 5, 4, 3, 2, 1, and of course 0 became the building blocks of modern mathematics. This sequence identified zero (zephirum) not only as “nothing” but as a number in its own right. Leonardo used this sequence to show merchants how to use these numbers in everyday transactions.
There are two features of this numbering that are helpful: one is the idea that the position of a number in a sequence indicates its size (so 90 is 10 times 9). The other is that this position system only works if one of the ten numerals stands for nothing. In other words, the language of mathematics only works if zero is also considered the sign for an operation—the process of changing a digit’s value by moving its place.
The Fibonacci numbers are Nature’s numbering system. They appear everywhere in Nature, from the leaf arrangement in plants to the pattern of the florets of a flower, the bracts of a pinecone, or the scales of a pineapple. The Fibonacci numbers are therefore applicable to the growth of every living thing, including a single cell, a grain of wheat, a hive of bees, and even all of mankind. Nature follows the Fibonacci numbers astonishingly. But very little of the beauty of nature. The Great poet Rabindranath Tagore also noted this. If we study the pattern of various natural things minutely, we observe that many of the natural things around us follow the Fibonacci numbers in real life, which creates a sense of strangeness among us. The study of nature is very important for the learners. Realizing that everything connects to everything else.
UNI-a prefix meaning having or consisting of only one.
VERSE-A single metrical line in a poetic composition; one line of poetry.
The main purpose of chlorophyll is to capture photons of energy. When growth is no longer a priority for a plant in nature, it will naturally sink its "no longer needed" nitrogen and switch pigments to carotene and anthocyanin, which are better-suited pigments for environmental protection rather than capturing photons (growth). This is because of all nutrients used by a plant, nitrogen in its nitrate form is highly stable and required for the rigours of electron capture, ammoniacal nitrogen readily evaporates into the air during cure/dry. The ripening stage is no longer about growth, but the main priority is protecting trichomes and opening up the oxidative capacity of the plant so it can focus 100% of filling those trichomes. Nitrogen in its elemental nitrate form is no longer required during ripening to the same degree as it was; the vast majority of growth is over. Understanding when to shift a plant's focus from rapid growth to maturation is the key to maximizing the trichome quality and terpene profile of the final product. That is the single most important decision you will make during a grow in terms of dictating smoothness, taste, and smell, I have found from my limited years of dedicated and driven study into the matter.
Synthetic nutrients maintain a near full nitrate ratio comparable to ammoniacal nitrogen, whereas organic nutrients maintain a supply of ammoniacal and convert only to nitrate what is needed. This is far more beneficial for organic growers when it comes to smoke smoothness, as the higher levels of ammoniacal nitrogen are far easier to dispose/convert than nitrates come harvest, cure/dry with far more efficient margins held within leaf/bud when it comes to conversion to aminos. A good dry and cure can go a long way to a quality smooth smoke, but without a proper senescence sequence or triggered autophagic sink of nitrogen, then you will be asking far more than can be expected from a dry/cure. It's one thing to sink the last 5-10% and convert the last drips to amino acids during the dry and cure, but leaving the plant fully charged with nitrogen in the form of nitrate is simply not going to taste good, no matter how much smoke you blow up your own ass or how long you have been growing. Means nothing. Smoke quality relies on pre-harvest physiology, not just the dry and cure! The dry and cure cannot fix poor pre-harvest metabolic states. A high-quality, smooth smoke requires correct signalling before harvest. While a proper dry and cure can gently soften the edges and convert the last 5-10% of sugars into amino acids, it fundamentally cannot perform magic on a fully fertilised plant. You cannot oxidise stable nitrates (+5 charge) into a smoother state during drying. Ultimately, accept it's going to be a heavy smoke and be grateful, onwards upwards!
The way in which ammoniacal nitrogen reacts to carbon sugars is critical in how it acts during oxidation and is responsible for oxidizing leftover sugars that can make the buds smoke harshly. Limited ammoniacal equals limited oxidation of excess carbons held within tissues. Just so you know, it's not an anecdotal rule that's unresolved and unsubstantiated. It's called the Maillard reaction. Signal triggers and stressors. Normally, you can only use 10-30% ammoniacal nitrogen, but as harvest closes, it is beneficial to increase this ratio 50% of ammoniacal as it assists in steering the plant towards maturation. Rather than running 50% ammoniacal nitrogen continuously until harvest, use it in short, targeted flushes/additions (e.g., 7 days) to slow internode elongation and hasten maturity. Use alongside forced larger "dry backs" (allowing the root zone to dry out significantly between watering events). Raising the Electrical Conductivity EC of the medium to create mild osmotic stress. Widening the day-to-night temperature difference and increasing the vapour pressure deficit VPD.
How would you like your cola's sir? Arm length, please, no gaps with trichomes as thick as snow.
2 likes
comments
Share
9
Week 9. Flowering
16d ago
1/23
101.6 cm
Height
12 hrs
Light Schedule
31 °C
Day Air Temp
6.4
pH
No Smell
Smell
55 %
Air Humidity
22 °C
Solution Temp
22 °C
Substrate Temp
23 °C
Night Air Temp
378.54 l
Pot Size
7.57 l
Watering Volume
1400 PPM
CO₂ Level
Ultraviolet_ We become aware at such times of man's wonderful ability to bring seemingly unrelated elements into harmonic balance, and receive a glimpse into a world where everything exists in conscious sympathetic attunement to everything else. When we listen to a piece of music that seems to strike a beautiful chord somewhere inside us, or view a painting that simply glows with harmonic awareness well executed, we probably do not spare much time to contemplate the wonderfully intricate combination of vibrations that our sensors make it possible for us to perceive. We tend to appreciate the relationship between the parts of something by an awareness of the harmony or dissonance of the whole. This ability enables us to say "what a beautiful house!" instead of "analysis has proven that this collection of building materials exhibits certain elements of harmonic proportion." While this ability to instinctively appreciate the beauty of true harmony has an important role to play in evolution, a more analytical understanding of the laws involved can be most useful. This is especially so if we wish to create works of art where each part exists in true harmony, not only with the other part of that particular whole, but with the universe within which the creator and the created exist.
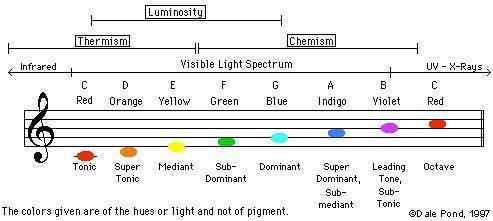
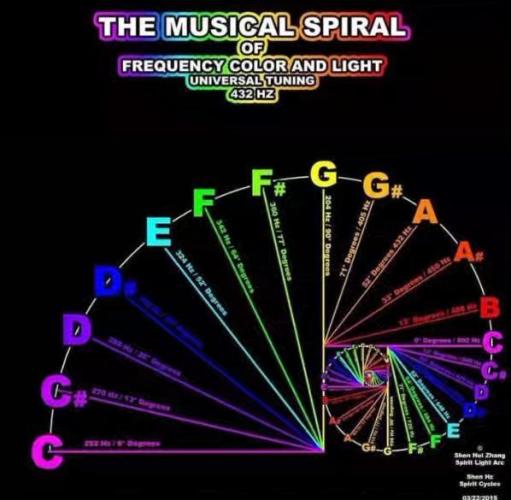
Everything vibrates. From the most dense matter to the most subtle cosmic rays, everything which our senses allow our thoughts to become aware of, can be specified in terms of wavelength or frequency of vibration. These two terms define the same thing, but from different points of view.
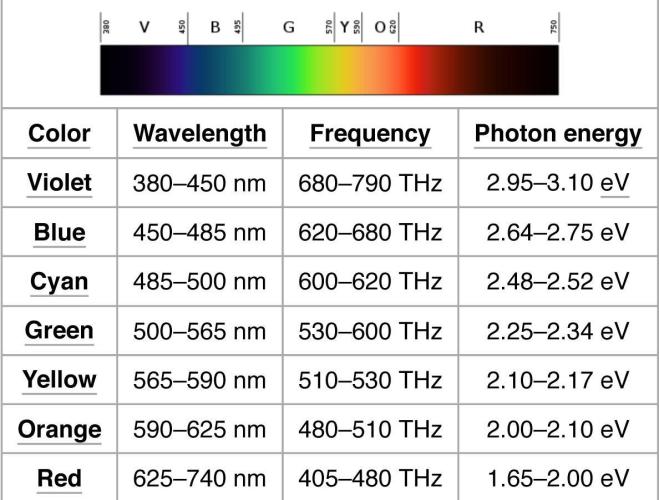
The following are the approximate wavelengths of various energy carriers:
Cosmic rays 0.000,000,001 mm
Gamma rays 0.000,000,1 mm
X rays 0.000,500 mm
Ultraviolet rays 0.003 mm
Visible light 0.006 mm
Infrared 0.01 mm
Sound waves 1 meter
Radio waves 300 meters
Violet light 400 nm to 450 nm
Blue light 450 nm to 500 nm
Green light 500 nm to 570 nm
Yellow light 570 nm to 590 nm
Orange light 590 nm to 610 nm
Red light 610 to 700 nm
1 nanometer (nm) = 0.000,000,1 cm = 1/10,000,000 cm
If a guitar string is plucked and we hear a sound, it is not too difficult for the human mind to associate this sound with the vibration of the guitar string. With color it is quite different. It is difficult for us to conceive that the color of a substance is not an inherent property of the substance itself, but an indication picked up by our senses of that substance's ability to absorb or reflect the light which happens to be shining on it at that moment. Neither the matter nor the light is colored. What happens is that the brain learns to differentiate between the frequencies reflected or transmitted by the substance the eyes are focused on. The same thing happens with sound. When we say "Oh! Listen, they're playing my favorite song," what we really mean is: "My brain has stored within it a particular pattern of frequencies. I have compared the new information being received with this stored pattern and have deduced the answer that the two patterns are similar within certain specified tolerances." The 'pleasure' involved could have something to do with our running the pre-recorded pattern at the same time, in 'sympathy' with the new pattern as it is received.
The word sympathy describes very well our ability to appreciate color and sound. It also describes the reason behind certain elements of harmony. For instance, if a substance vibrating at 100 cycles per second (tone 1) is in the proximity of another substance vibrating at 200 cycles per second (tone 2), we could perceive, if we had the right equipment, a certain sympathetic relationship between the two.
We will see from this that there is a uniform doubling of the first tone seen in the second. At various points along the waves, the two are the same in amplitude. At other points, they are at opposite poles to each other. This doubled frequency has more points of similarity to the original than any other frequency except the original itself. If the equipment we had available for measuring these two frequencies was a soundboard amplifier and a pair of ears, then we would hear what would sound to us like one tone. If we had the opportunity to hear one at a time, we would hear that although they sound the same, one is higher in pitch than the other. This characteristic of 'the same but different in pitch', musicians have called the octave. Any two tones produced where one has exactly doubled the frequency of the other is called an octave. Speaking in ratios, an octave would appear then as the ratio 2:1 or 1:2, depending on whether we are talking of an octave up or down.
A single note produced by almost any instrument will contain more than one wavelength or frequency. It will have a dominant frequency, the wavelength of which we would call the note's 'fundamental' or 1st harmonic. It will also have a varying number of upper harmonics, gradually fading in intensity into infinity or silence.
Natural harmonics always have the same pattern of intervals between them. The interval between the 1st and 2nd harmonic is a perfect octave; between the 2nd and 3rd a perfect fifth; between the 3rd and 4th a perfect 4th; and so on, the intervals becoming smaller and smaller until they lose any relationship with the western 12 tone scale as it exists at the moment. Just as an octave has certain elements of sympathy with its fundamental, so some intervals have been noted to be more perfectly in sympathy with the fundamental than others. The ratio of the 'perfect 5th' or interval of 7 semitones, as it occurs in the harmonic series, is 3:2 or 2:3, while that of the 'perfect fourth" is 4:3 or 3:4. All the tones in the western 12 tone scale can be expressed in terms of the ratio between the upper tone and its fundamental. This would seem to be an ideal way of generating a scale from any given fundamental and several attempts have been made to do this, the Pythagorean system being probably the most well known. Although when working with a single tone instrument playing on its own, the Pythagorean formula works wonderfully well, if we had several instruments tuned this way together and asked them to play almost any western music, we would find that at times they sounded quite out of tune to each other.
The lack of flexibility of the various scale systems based on the harmonic series has led to what is known as the 'tempered' scale. This uses as its primary unit of interval the ratio of the octave or 2:1. It then proceeds to divide the interval between any fundamental and its upper octave into 12 smaller intervals by applying the ratio: two to the one-twelfth power, to one (21/12:1). This equals 1.059463094, so by multiplying any frequency by this number, we will obtain the tempered semitone next up from our fundamental. We will also find that any tone twelve semitones up from any other tone, in a scale generated in this way, will have exactly double the frequency.
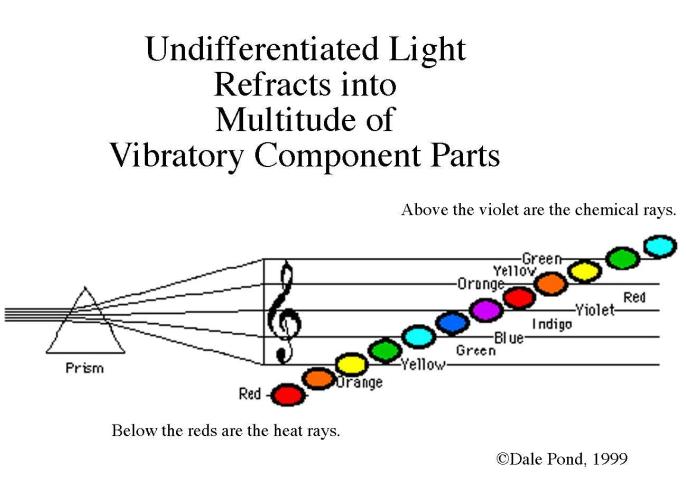
If we took the note middle C on a piano and halved the wavelength, we would have the note C one octave above. If we halved this, we would have the C above, and so on. However, within about 6 octaves, we would find that although a 'sound' was being produced, no human ear could perceive it. If we kept on going, halving and producing upper octaves of our fundamental C, we would proceed through the infrared band, into the visible light spectrum. If we happened to be outside during the day, we would, for one octave only, see the note C with our eyes. The next octave above would already be in the ultraviolet band, and outside the eye's sensitivity range. If we can think of color as being an indication of a substance's vibratory rate or wavelength, we may begin to see a relationship that could exist between the color and sound spectrums.
The logical extension of what has so far been said is that there exists a scale in the color spectrum that corresponds exactly to the scale in the sound spectrum, each color tone being an octave of the equivalent note in the sound range.
This is not the end of the story but only the beginning. If we can for the moment accept that any wavelength in one band has upper and lower octave stretching out to infinity, then the next question is 'fine, but what shall we use as our fundamental? A particular color? A particular sound frequency?' The musicians among us will probably say 'A 440'. This means that the note A should vibrate at 440 Hertz, or 440 times per second. They would tell us that this is standard pitch has been adopted by most orchestras around the world; pianos are tuned to it, instruments are constructed to formulas based on it, and so to them it would probably seem the most appropriate place to begin. Some of these musicians may know of the battle that is still raging with regards to this being the standard, but few would know why A = 440 Hz was chosen except that it worked when it was set to be a suitable compromise between the many different pitches in use at the time.
432 Hz is said to be mathematically consistent with the patterns of the universe. Studies reveal that 432 Hz tuning vibrates with the universe’s golden mean PHI and unifies the properties of light, time, space, matter, gravity, and magnetism with biology, the DNA code, and consciousness. When our atoms and DNA start to resonate in harmony with the spiraling pattern of nature, our sense of connection to nature is said to be magnified.
Another interesting factor to consider is that the A=432 Hz tuning correlates with the color spectrum, while the A=440 Hz is off. Audiophiles have also stated that A = 432 Hz music seems to be non-local and can fill an entire room, whereas A=440 Hz can be perceived as directional or linear in sound propagation.
Once you adopt the idea that sound (or vibration in general) can have an equalizing and harmonizing effect (as well as a disturbing effect), the science of harmony can be applied to bring greater harmony into one's life or tune to specific energies. There is a form of absolute and relative harmony. Absolute harmony can, for example, be determined by the tuning of an instrument. The ancients tuned their instruments at an A of 432 Hz instead of 440 Hz - and for a good reason. There are plenty of music examples on the internet that you can listen to in order to establish the difference for yourself.
Attuning the instrument to 432 Hz results in a more relaxing sound, while 440 Hz slightly tenses up the body. This is because 440 Hz is out of tune with both macro and micro cosmos. On the contrary, 432 Hz is in tune. To give an example of how this is manifested microcosmically: our breath (0,3 Hz) and our pulse (1,2 Hz) relate to the frequency of the lower octave of an A of 432 Hz (108 Hz) as 1:360 and 1:90.
It is interesting to note that 432 Hz was the standard pitch of many old instruments and that it was only recently (19th and 20th century) the standard pitch was increased. This was done in order to be able to play for bigger audiences. Bigger audiences (more bodies) absorb more of the lower frequencies, so the higher pitch was more likely to “cut through”. One of the oldest instruments of the world is the bell ensemble of Yi Zeng (dated 423 BC), tuned to a standard F4 of 345 Hz, which gives an A= 432 Hz. The frequency of 345 Hz is that of the platonic year! Similarly, many old organs are tuned in A=432 as well; for example, St. Peter’s Capella Gregoriana, St. Peter’s Capella Giulia, S. Maria Maggiore in Rome.
Maria Renold’s book “Intervals Scales Tones and the Concert Pitch C=128 Hz” claims conclusive evidence that 440 Hz and raising concert pitch above scientific “C” Prime=128 Hz (Concert A=432 Hz) disassociates the connection of consciousness to the body and creates anti-social conditions in humanity.
The difference between concert pitch A=440 Hz and Concert A=432 Hz is only 8 cycles per second, but it is a perceptible difference of awareness in the human consciousness experience of the dream we share called existence.