
Likes
Comments
Share


@ojulhao
Follow
[Germination: 07/06 - 13/06]
For germination, I followed a scientific article (https://pubmed.ncbi.nlm.nih.gov/33732764/) that describes a method with a high success rate. Here it is:1. Prepare a 1% hydrogen peroxide solution.
2. Leave the seed in the solution for 18–24 hours, no longer.
3. Use the normal paper towel method.
Do this in a clean environment, avoid touching the seeds directly, use gloves if possible, and clean everything.
I started the process on the 7th of June 2026, with 5 seeds. After three days, only 3 germinated, and I placed them in a Jiffy Pellet.

Likes
35
Share


@Ganjagrandaddy
Follow
Day 49 today and the start of an attempt at drought stressing her . The science says to withhold water until a noticable droop on their frame bit rhisnlady is a solid limbed , woody beast so that won't happen the same at all. To counter this I expect around 7 days is all I will push it too so right on point for a final weeks feeding before harvest and ripening. The effect hoped for a is a noticable difference in the thc and oils produced. It sounds similar in move as other methods like stem splitting , taking chunks out as though being threatened by wildlife ect.. but is more in keeping with seasonal changes and auxin changes. I did leave my last crop in the dark for over 7 days drying but didn't benefit too much from that. It is something I have wondered about since reading about it.
I pulled the Red Hot Cookie and the Sweet Afghani Delicious as rhe tank took a turn for the worse so took the sensible option. S.a.d is drying but I quick dried the Rhc and had a few nice joints from her. not her full potential but certainly helped me sleep. lol
Amnesia haze is mid way through her second pistil flush with plenty of fresh ones backing them up too . Her overall size and bud yield should be even better once they begin to discolour and push even more trichromes from this amazingly sweet and fruity girl.
She was the 1st to show hunger issues before i used the megacrop addition so was taking all her stashed nutes . shebhas rewarded me with a very good sized plant that is a star to work with.
He buds are rock solid to touch and leave a beautiful smell on my fingers when touched. The trichromes are slowly being hidden by the newer pistil growth but inexpectbsjebwill build on them too. I didnget a day of high humidity when my dehumidifier filled up and switched off. This managed to allow some bud rot onto 1 cola but I caught it early enough to not be an issue now. She has done her breeders proud indeed. I hope the pics do her justice as I love this lady and how's he is finishing. 2 weeks max now I think. Enjoy the vid and pics. thanks for stopping in. Roll on the sweet seeds autos.....

Likes
15
Share


@Style_weed
Follow
Dopo aver raccolto la piu piccola che era la più matura ho iniziato il lavaggia di altre 3 mentre a 1 inizierò dalla prossima volta... I fiori più maturi sono bellissimi e il profumo è altrettanto fantastico!!! Le annaffierò altre 3 volte con sola acqua e siamo al traguardo!!!

Likes
33
Share


@Philhsy
Follow
Removed powerbuds, caused ph fluctuations, but daaamn this grow is insane. Will be fun to se final weight!
Been fighting this auto dwc for a while now. All green all the way, super nice terps menthol tones love it 🤩 soooon harvest, waiting for bit more trichromes and maturing,
Likes
15
Share


@SweetheartOG
Follow
The girls are fully in flower this week which of course is a GREAT sight to see when opening the tent. The smell in the tent is so sweet and fruity it's already making your mouth water!! So this week I feed the girls with a little of the living soil veg so they are still getting a helping hand. They were also given the living soil bloom and some barley which I will admit is the first time using it but so far me plus the girls are very much enjoying!! I have done the necessary light de-foliating on all 4 girls over the past 2 weeks and more LST. Happy toking guys and girls!! Will keep you all posted with more videos through the week so please keep checking back 🙂

Likes
2
Share




Likes
14
Share


@ao_grows1978
Follow
The OG Kush made it through her final week of flower...56 days in total. Super sticky and smells like a mix of limonene and pinene and a hint of sweetness on the back end. I'm definitely happy I have a couple more of these beans. Super smooth run, chunky resinous buds. Thank you to everyone that visited to check my first Grow Diary out. You made it a really cool experience ✌️☺️.
https://www.instagram.com/ao.grows?igsh=MXR1dWZxMWhlNnF6bA==
Check out my Instagram for clips I made for this grow.

Likes
14
Share


@Hempface86
Follow
All right guys run day 33 in the middle of week four and this is my sixth time trying to upload this diary so fingers crossed this goes through... As you can see so far so good this is my first run with organics so these plants are nowhere near as big as my plans typically are so don't think it's due to genetics it's because I'm not giving them the cocktail mix that I give or gave my older ladies but I will say I'm probably going to go back to that because I like huge plants and I cannot lie 🤥 the taste of the flower I grow with synthetics is just as good if not better than what I'm getting at the Dispensary I believe it all boils down to how you dry it and cure it.. with that being said shout out to Sensible Seeds for giving me an opportunity to grow their genetics and give him a shot and so far so good.. I hope everybody is doing well have a great day and happy growing ✌️

Likes
5
Share


@CannDy420
Follow
Die Pflanzen haben sich an die Neuen töpfe gewöhnt und wachsen gut an nächste Woche sind sie denk ich ready für die erste Nährstoff Zugabe .
Likes
7
Share


@Aleks555
Follow
We are entering the third week of growth for our FBA 2503 strain from 42Fast Buds, and everything is progressing beautifully. We've recently transplanted the plant into a 10-liter pot, giving it more room to thrive. Even though it's an autoflowering strain, we've decided to experiment with a 12/12 light cycle—12 hours of light during the day and 12 hours at night—to see how it responds.
The daytime temperature peaks at 30°C, with an average range between 27°C and 28°C, while humidity is consistently maintained at 60% to 65%. The plant has reached a height of 14-20 cm and is already showing three sets of leaves. We're preparing to start low-stress training (LST) to help guide its growth.
In addition, we've begun feeding our beauty with nutrients from Xpert Nutrients, and we’re really pleased with the results so far. The plant looks strong and vibrant, and we're excited to see how it develops over the coming weeks!



Likes
11
Share
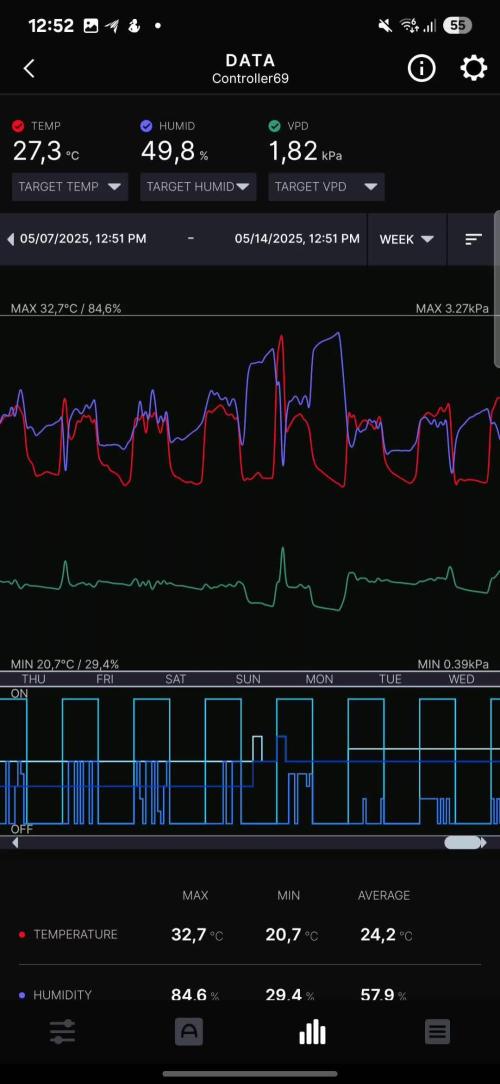

@WeedWagon
Follow
Day 30 - 37:
- first week of flower
- no signs of flower yet
- Day 36 Transplantation into 15 l Pots
- lst all the way (could be better every 2-3 days)
- 2l every 2-3
Likes
67
Share


@Mr_und_Ms_Lue_Roots
Follow
Hallo und herzlich Willkommen,
Viel gibt es hier momentan nicht zu sagen. Die eine Dame Stagniert im Wachstum und benötigt dahingehend immer weniger zu trinken. Die andere hingegen benötigt jetzt einiges um sie wieder auf den richtigen Pfad zu bekommen. Mal schauenwie sie nächste Woche wirkt.
Ansonsten allen Growmies eine angenehme und erfolgreiche Woche gewünscht!
Vielen Dank im Voraus für das verfolgen oder den Besuch dieses Update, Kommentare wie auch likes!
VG✌️

Likes
13
Share


@BigDaddyK
Follow
I plan to grow the best 2 in hydro and the best 2 in soil
On 40% power
Big Thank you to Medicgrow for the opportunity to use the LED - I am under no pressure or obligation to promote this item , I will give you my honest opinion, and we will see the results , it’s actually a bit bigger than imagined , first test with the lux meter showed awesome spread and distribution, I have one thing that I didn’t understand, using the hangers , you need a degree in engineering to understand , I looked , I pondered, I scratched my head a little smoked a joint , still couldn’t comprehend,so I used 6 ratchets = 🤪
All of them are cuttings I took from a naughty strawberry cheesecake
Ps The big green power supply there’s nothing to do with the medicgrow fold 6 ,( the power supply is in the middle )it’s from another light that I just can’t be bothered to move because I’ve got nowhere to put it
Wednesday-watered the soil , topped up the pots and added 2ml of A+B hydrogrow
Thursday upped the power to 60% this is the recommended setting , but I was a little cautious, I didn’t want to burn anything
Probably going to cull down to 6 this week , 3 of each - throw the others - see what happens


Likes
27
Share


@Bread_n_Buds
Follow
Our Zamnesia photoperiod plants have also started their journey into the magical flowering period, this is the first week.
So born on September 1st, on October 1st they went into flowering and these are the photos of the first week, when the ladies begin to understand that they have to do their magic.
---- The general environmental conditions are good, the heat has increased a little more but for the first weeks of flowering it is not a big problem, let's remember to never exceed 27 degrees centigrade in the last 4 weeks to avoid compromising the quality of the flowers. If necessary, open the growbox but never let it exceed 27-30 degrees
where 30 is already a lot. The positive thing is that as soon as the lights go out the change is already a little visible and as autumn progresses we will also improve. The humidity is fluctuating but we try to control it with two dehumidifiers when necessary, pushing both of them possibly not into the growbox but directly into the room. (I dehumidify the room and the air in the growbox should be dehumidified too)
--- We are at a top secret level here so I can tell you that it is called XXXX Test 2 and we really like how they are growing. They were practically identical so I decided to top one and let it grow like a bush keeping some branches underneath but not in main lining style. The other one is growing straight and beautiful as the sun. Very wide internodal distance, wide plant, definitely to be supported soon.
--- Fertilization schedule - The fertilizers are always Plagron and I am following the table that I generated on the site depending on the chosen substrate. Create your own card and follow it, never be presumptuous my friend as I have been in the past, follow the card and look at the plants, not all need the same amount of fertilizer. We have come to the time to give a good dose of iron in the first weeks of flowering, already of race life not sprayed but mixed with fertilizers. Alga Bloom is now running instead than alga grow
---- https://plagron.com/en
- Power Roots - 1ml/l
- Alga Bloom - 4ml/l
- Pure Zym - 1ml/l
- Sugar Royal - 1ml/l
- Vita Race - 5ml/l
--- Dehumidifier now runs between 50% and 55%
--- Plagron 100% Organic pack can be found on Zamnesia at: https://www.zamnesia.io/en/11457 -pla gron-easy-pack-natural.html
--- new strain coming soon no description of the strain because it is a secret
- Fully organic soil and nutrients -- https://plagron.com/en buy on www.zamnesia.io
- air system --- https://www.secretjardin.com/
- Growbox & Light —— www.Marshydro.eu
- Music and sound --- I made my girls listen to 432hz frequencies and music from www.radionula.com
- Z --- You can find these seeds, much more from the world of cannabis, mushrooms and an incredible series of accessories and gadgets on the reference site not only mine but of many growers
—— https://www .zamnesia.io
Likes
37
Share


@Wrongholn519
Follow
Edit: I came home from work after making my weekly log entry yesterday and I could smell my grow from my driveway... so I asked for help. THANK YOU OOZLEFINCH FOR THE HELP.. THIS IS WHY WE ASK FOR HELP...
I left a question on here stating that my new canister wasn't working and asked for generic advise. He left an answer that pointed out the fact that my exhaust fan might be set waaaay to high... and it was! It turns out there's a limit to the amount of air that filter fan 'scrub', and my exhaust fan goes way over that limit even on 6 or 7.. I turned it down to 3 and the smell got better immediately. ☮️
Still on auto pilot at this point... I'm really close to harvest.. and I'm not even on day 60 😲. I created videos this weeks, they capture the plants in ways that a still picture just won't.
My entire house stinks.. my neighbors are looking at my funny, and my wife thinks a family of skunks is hiding in our walls... I've never had this problem.. I even got a brand new carbon air scrubber and its not helping 🤣
Process/ Environment changes since last week:
- To help preserve the terpenes I've been trying to lower my temperature, which is hard without an AC unit I'm not plugging in an ac just to grow some plants. I'm at 24- 26 degrees C (75 - 82 F).
- I've been lowering my humidity accordingly to keep VPD in a good range
- I lowered my lights closer to my plants, turning them up would just add to the heat.
- I'm defoliating as much large fan leaves off as possible
- My watering pH is at around 6.7 to 6.8.. I will not be flushing these plants
- I rotate my plants every day to make sure I'm getting light to hit every node it can
This week I'm pulling out the digital scope and doing the first trichome checks for these babies. The cheese auto is going to get the chop by the end of next week.. I hope you like the video I added of it.. it's the easiest plant I've ever grown and she's going to be FIRE.
The blackberry auto is giving off a gas/berry smell and is very sticky to the touch. A beautiful triploid... my first and I hope to get many more.
The Forbidden Runtz smells just like a zkittles does, it's bang on. Very dense very sticky buds. I almost tossed her during the seedling stage too due to her mutation. I'm glad I reconsidered ☮️
Thank you Fastbuds
Likes
8
Share


@Hash268
Follow
As we approach the end of her cycle as I flush she is just stinking up this place like really amazing, the moon rock buds a super dense and hard I try not to touch her but trust me I'm pleased stay tuned for the harvest 😎.

Likes
38
Share


@Zurban_Poison
Follow
amazing variety! struck me during the growing season with streaming growth! and of course in bloom! She did not stop growing for a second throughout her life.)) From the beginning in height and then in width)) Well, what else can I say? you see everything.

Processing
Likes
2
Share



Likes
10
Share


@SkunkyDog420
Follow
Hallo zusammen 🤙.
Sie wächst sehr schön und macht keine Probleme.
Rabattcode für den BIOTABS-Webshop https://biotabs.nl/en/shop/ GDBT420, damit erhalten Sie 15 Prozent